

首款脑胶质瘤溶瘤病毒产品正式上市
发布时间:2021-11-12 11:52:04 | 阅读:次| 关键词:
- [案例]INC国际Rutka教授团队发表成果助力儿童胶质瘤免疫治疗
- [案例]对抗婴儿痉挛引发的癫痫治疗案例
- [案例]6岁男孩脑干胶质瘤 “巴爷爷”成功手术后都可以包水饺啦
- [案例]得了这种胶质瘤,竟有望通过全切手术治愈!
- [案例]【治疗指南】INC国际福洛里希、Schroeder教授参与欧洲前床
- [案例]脑内血流搭上“直通车”该怎么办?Lawton教授7步拿下胼胝
2021年11月1日,日本一三共宣布其溶瘤病毒疗法Delytact正式在日本上市,用于治疗恶性胶质瘤患者,定价143万日元(折合人民币8万元)一针。这是一款获批用于原发性脑胶质瘤治疗的溶瘤病毒产品。(参考阅读:快讯|脑胶质瘤溶瘤病毒疗法获批上市)
据了解,Delytact(Teserpaturev/G47∆)是一种基因工程溶瘤单纯疱疹病毒1型(HSV-1),是一三共与日本东京大学医学科学研究所的联合开发的三代溶瘤病毒。
根据相关临床数据统计,溶瘤病毒疗法Delytact治疗恶性脑胶质瘤的1年生存率高达92.3%,是常规治疗的6倍左右,提升了恶性脑胶质瘤患者的生存率。
溶瘤病毒是什么?
溶瘤病毒(oncolytic virus,OV)是一类能够特异性复制并引起癌细胞凋亡,同时保留正常组织不被损害的病毒。溶瘤病毒疗法是一种前景广阔的癌症治疗方法,迄今已报告了70多项临床试验。OV可以分为2个主要类别——天然病毒和转基因病毒。天然存在的OV包括呼肠孤病毒、新城疫病毒(NDV)、肠病毒和麻疹病毒(MV)等,而大多数OV都经过基因修饰,以增加病毒向肿瘤细胞的向性、提高病毒的选择性复制和裂解潜能,并增强宿主的抗肿瘤免疫。
具有溶瘤活性的病毒有多种类型,下面展开详细介绍。
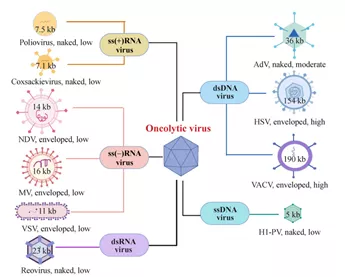
图1一些用于癌症治疗的溶瘤病毒的示意图
许多病毒已被用于癌症治疗,包括双链DNA病毒,如AdV、单纯疱疹病毒(HSV)和VACV;单链DNA病毒,如细小病毒;负义单链RNA病毒,如新城疫病毒(NDV)、麻疹病毒(MV)和水泡性口炎病毒(VSV);正链单链RNA病毒,如脊髓灰质炎病毒和柯萨奇病毒;和双链RNA病毒,如呼肠孤病毒。naked/enveloped描述病毒类型;high/moderate/low描述OV的基因组容量。
腺病毒
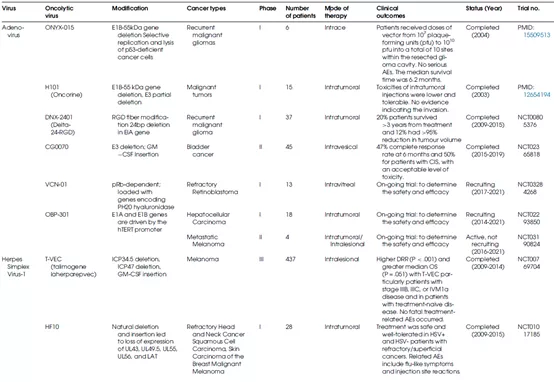
腺病毒(Ads)是一种非包膜的双链DNA病毒,具有30-38 kb的线性基因组和二十面体衣壳。感染人的Ads根据其DNA同源性、致癌性、血凝作用和血清中和特性可分为7种(A-G)。基于对腺病毒基因组结构和生命周期的了解以及腺病毒稳定的生理特性,对腺病毒进行基因操作相对容易实现。大多数基因工程溶瘤腺病毒(OAd)来源于Ad血清型5(图2),HAdV-C5是较常研究的一种。
在必需的腺病毒基因中进行小部分的缺失并进行遗传修饰可诱导溶瘤腺病毒的肿瘤选择性(图3)。E1B-55K和E1A基因的缺失可分别引起p53和视网膜母细胞瘤(pRb)突变癌细胞中Ads的选择性复制。溶瘤腺病毒的先驱是ONYX-015,该病毒在E1B-55K基因以及E3B和rid基因中有14.7k基因缺陷。Oncorine(H101)是一个被监管机构批准用于临床的OV,与ONYX-015结构相似,它删除了E1B55K蛋白在正常细胞中使p53失活的编码区,导致该病毒只能在p53功能异常的恶性细胞中复制;除了E1B编码基因的缺失外,Oncorine还在E3区域进行了部分缺失。E1A区域的缺失也被用来构建溶瘤Ads,由于E1A与pRb结合,E1A中24bp(Delta24)的缺失导致E2F的释放,较终导致肿瘤细胞中的病毒复制。2005年,Oncorine(H101)被中国FDA批准用于联合顺铂或5-氟尿嘧啶(5-FU)或两者联合治疗鼻咽癌。其他正在进行临床评估的OAd包括LOAd703、VCN-01、Telomelysin(OBP-301)和ONCOS-102,这些OAd产生了合适的抗肿瘤功效并且在患者中具有良好的耐受性。
图2野生型血清5型腺病毒基因组
基因组由4个早期转录元件(E1、E2、E3和E4)和5个与腺病毒颗粒组装相关的晚期表达基因(L1-L5)组成。E1基因是激活早期基因转录所必需的;病毒DNA复制需要E2基因;E3基因是调节和逃避宿主免疫反应、通过细胞凋亡防止细胞过早死亡以及新粒子组装完成后合适细胞裂解所必需;E4基因参与病毒RNA代谢和运输、宿主细胞蛋白质合成的优先下调以及病毒DNA复制的增强。E1和E3基因的缺失较多可容纳7.5 kb的外源DNA,通常用于基因治疗。E4基因缺失进一步减少了载体特异性免疫反应的诱导,并较大限度地减少了包装细胞系中具有复制能力的病毒的生长。
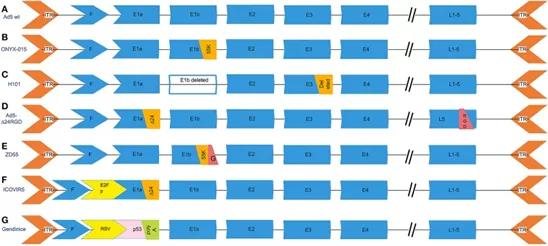
图3常见溶瘤腺病毒载体的遗传修饰
(A)野生型血清5型腺病毒。(B)ONYX-015,删除了能够在p53缺陷型人类肿瘤细胞中复制的E1b-55k(2496-3323)区域。(C)H101,删除整个E1B基因和E3区域中的78.3-85.8 nm基因片段,该片段负责腺病毒死亡蛋白的编码。(D)Ad5-Δ24RGD在E1A区域携带24 bp(919-943)缺失,负责结合Rb蛋白和RGD基序插入纤维。(E)ZD55删除了E1B 55-kD基因并带有外源基因。(F)ICOVIR5,溶瘤腺病毒,其中内源性E1A启动子已被用DM-1绝缘的人E2F-1启动子取代。(G)Gendicine是一种重组人血清型5腺病毒,其中E1区被人野生型p53表达盒取代。Gendicine是一种重组人血清型5腺病毒,其中E1区被人野生型p53表达盒取代。p53基因由带有牛生长激素(BGH)poly(A)尾的劳斯肉瘤病毒(RSV)启动子驱动。
单纯疱疹病毒
单纯疱疹病毒(HSV),是1型HSV(HSV-1),是作为OV研究较广泛的DNA病毒之一。HSV-1是一种有包膜的双链线性DNA病毒,具有152 kb的大基因组,其中部分基因对复制来说是不必要的,因此在不限制病毒包装效率的前提下,为添加工程化转基因留下了空间(图4)。HSV-1强大的毒力和免疫原性具有“双刃剑”效应:毒力和免疫原性使合适的溶瘤和抗肿瘤免疫反应成为可能;然而,其快速免疫介导的病毒清除和潜在的细胞毒性会引发递送和顺利问题。为了实现肿瘤选择性和优化疗效,科研人员已经创建了几种HSV突变体(图5)并将其应用于多种类型的肿瘤。
T-Vec(talimogenelaherparepvec)是美国食品和药物管理局(FDA)批准的一个OV,是一种溶瘤性HSV-1,其基因组中有2个缺失:ICP34.5和ICP47。编码神经毒力因子的ICP34.5缺失阻止了病毒在神经元中的复制,而不影响病毒在其他细胞,是肿瘤细胞中的复制。ICP34.5对于阻断宿主抗病毒先天免疫蛋白激酶R-干扰素(PKR-IFN)途径至关重要;大多数肿瘤细胞缺乏PKR通路,使ICP34.5缺失的HSV对癌细胞更具选择性。T-VEC含有GM-CSF(代替ICP34.5),可促进树突细胞成熟并增强对肿瘤细胞的免疫反应。编码抗原呈递控制剂的ICP47的缺失导致肿瘤相关抗原和病毒抗原进入主要组织相容性复合体(MHC)Ⅰ类复合体,并促进针对肿瘤细胞的免疫应答。
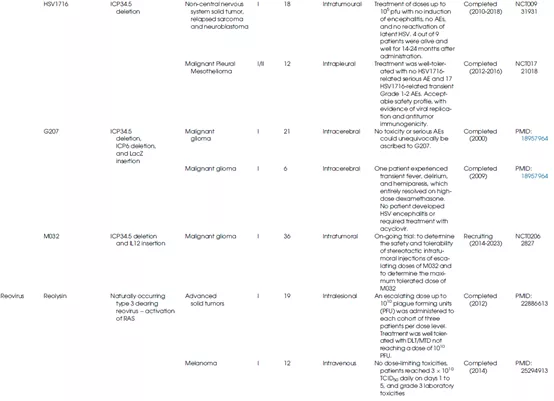
另一种HSV突变体HF10也在几种肿瘤中进行了临床评估。较近,一项研究报告在小儿癌症患者中静脉施用HSV突变体HSV1716与肿瘤内施用相比,全身递送显示出更好的耐受性,同时确定了相似的临床反应。
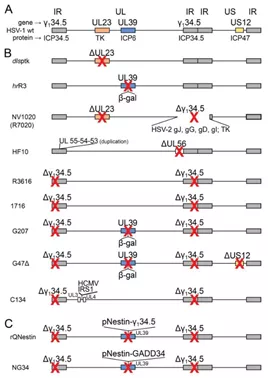
图4野生型和重组型HSV基因组示意图
(A)野生型HSV:IR,inverted repeats;UL:unique long;US:unique short.(B)具有单个和多个突变的溶瘤HSV。(C)转录靶向型溶瘤HSV。与肿瘤特异性相关的基因显示为彩色框,其名称显示在基因组线上方;基因产物的名称显示在基因组线下方。红色叉号表示通过删除(Δ)或插入导致基因的失活。
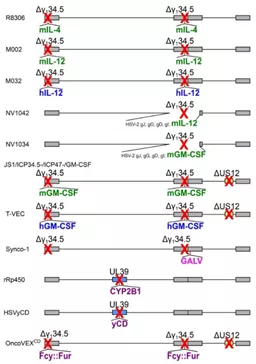
图5基因武装型HSV基因组示意图
绿色为鼠类细胞因子,蓝色为人类细胞因子,粉红色为异源融合蛋白,紫色为前药转化酶。红色叉号表示通过删除(Δ)或插入异源武装基因导致基因失活。
痘苗病毒
痘苗病毒(VV)是一种双链DNA病毒,基因组为180-200 kb,可在宿主细胞的细胞质中复制,具有感染细胞范围广、对肿瘤细胞的趋向性强、可携带大量外源DNA序列的能力等优势,可作为OV的研究候选。
为了提高痘苗病毒的选择性复制和裂解能力,已经进行了的外源修饰包括病毒胸苷激酶(TK)、痘苗Ⅰ型干扰素结合蛋白(B18R)或痘苗生长因子(VGF)的缺失。TK基因缺失使得痘苗病毒复制不依赖于宿主细胞TK的表达(图6)。研究表明,在正常细胞的细胞周期中,受E2F转录因子调控的TK的较高水平处于S期,但在癌细胞的整个细胞周期中其表达水平仍然很高。因此,痘苗病毒依赖宿主细胞核苷酸和TK活性的补偿,TK缺失的痘苗主要在肿瘤细胞中复制。B18R对Ⅰ型IFN的高亲和力可引发Ⅰ型IFN信号的阻断和痘苗病毒对健康细胞的感染。因此,通过B18R的缺失,健康细胞由于完整的Ⅰ型IFN应答而对痘苗病毒感染不耐受,而癌细胞由于Ⅰ型IFN通路的中断,易受痘苗病毒感染并较终溶解。VGF是一种表皮生长因子(EGF)类似物,通过与宿主细胞上的EGF受体(EGFR)结合激活RAS-MEK-ERK信号通路。因此,VGF的缺失导致病毒在EGFR-RAS信号异常的细胞(如癌细胞)中选择性复制。
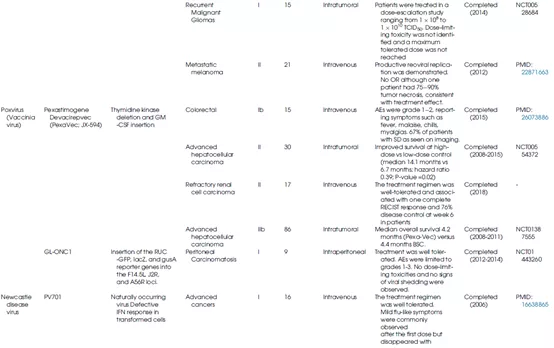
一种溶瘤痘苗病毒GLV-1h68(在临床试验中也称为GL-ONC1)是通过用3个编码β-半乳糖苷酶、β-葡萄糖醛酸酶和肾素荧光素酶/绿色荧光(RLuc-GFP)融合蛋白的表达盒分别替换病毒TK、血凝素和F145L基因。2012年至2014年间,在头颈部癌症患者中进行了静脉注射GL-ONC1的I期临床试验。30个月的随访显示,GL-ONC1联合标准化疗可提高总体生存率,这项研究还证明了病毒的顺利性。此外,由于标记基因(包括Ruc-GFP、β-半乳糖苷酶和β-葡萄糖醛酸酶)的表达,GL-ONC1还可实时监测细胞裂解和肿瘤治疗的情况。
Lister VV菌株的复制、细胞毒性和转基因能力不受缺氧环境的影响。其突变体Pexa-Vec(pexastimogenedevacirepvec,也称为JX-594)可阻断肿瘤血管生成,已进入临床试验(NCT01469611和NCT00554372)。JX-594在基因组上进行了3处修饰:插入GM-CSF编码基因以诱导系统免疫反应,删除TK基因以获得肿瘤选择性,以及在p7.5启动子控制下引入lac-Z基因。Park等人证明,JX-594静脉给药对难治性结直肠癌患者具有可接受的顺利性。
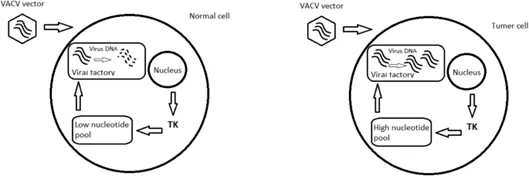
图6 TK与VV复制
TK的表达在正常细胞中普遍降低,但在快速增殖的肿瘤细胞中升高。TK缺失的痘苗病毒可以选择性地感染肿瘤T细胞,因为它具有高核苷酸库并开始DNA复制。然而,在大多数正常细胞中,TK基因的缺失大大降低了病毒的复制能力,甚至不能使病毒复制。
呼肠孤病毒
这种无包膜RNA病毒包含9-12段线性双链RNA。已有报道表明呼肠孤病毒在缺氧TME中能够维持其复制和溶瘤能力,并且在感染期间下调缺氧诱导因子1α(HIF-1型α)的表达。此外,呼肠孤病毒通过静脉注射,可以利用外周血单核细胞(PBMCs)和树突状细胞(DCs)作为细胞载体来逃避介导合适病毒清除的中和抗体(NAbs),这种生物学特性确定了其全身递送的合适性(图12)。
Reolysin(也称为pelareorep)是一种天然存在的呼肠孤病毒3型Dearing毒株,已在KRAS激活的恶性肿瘤中显示出强大的细胞毒性;其与化学疗法、放射疗法和免疫疗法的组合已在各种临床试验中得到评估。
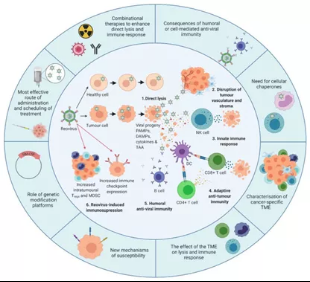
图12呼肠孤病毒的作用机制和发展概况
内圈说明了目前已知的呼肠孤病毒。1.在健康细胞中,抗病毒免疫反应限制呼肠孤病毒的复制,防止被杀伤。相反,致癌信号通路使肿瘤细胞易受呼肠孤病毒复制和直接溶瘤。2.呼肠孤病毒在肿瘤血管系统和间质中复制是由于细胞之间的相互作用改变抗病毒信号。3.肿瘤细胞的感染导致病毒子代、细胞因子和肿瘤相关抗原(TAAs)的释放,从而启动天然抗肿瘤免疫,包括细胞因子介导的旁观者杀伤和自然杀伤(NK)细胞介导的细胞毒性。4.树突状细胞(DC)吞噬TAAs并将其呈递给CD4+和CD8+T细胞后产生适应性抗肿瘤免疫,这有助于启动肿瘤特异性细胞毒性T淋巴细胞(CTL)。5.除了先天性和适应性抗肿瘤免疫反应外,还可诱导体液抗病毒免疫,从而产生呼肠孤病毒特异性中和抗体(NAbs)。在诱导抗肿瘤/抗病毒免疫应答后,调节性免疫机制被“开启”以控制持续的免疫应答,包括免疫检查点的上调和调节性T细胞(Tregs)和/或髓源性控制细胞(MDSCs)水平的增加。
外圈突出了提高呼肠孤病毒的疗效的重点研究领域,包括进一步了解:(i)体液和/或细胞介导的抗病毒免疫对呼肠孤病毒疗效的影响,这将为细胞伴侣的发展或需求提供信息。(ii)肿瘤微环境(TME)及其如何影响呼肠孤病毒溶瘤和抗肿瘤免疫。(iii)呼肠孤病毒用于直接溶瘤的细胞决定因素,包括呼肠孤病毒抗性机制。(iv)转基因呼肠孤病毒平台的潜在益处。(v)呼肠孤病毒计划,以较大限度地提高病毒的交付和效力,包括较佳的病毒管理途径。(vi)旨在促进直接溶瘤和抗肿瘤免疫反应的组合方法。
麻疹病毒
麻疹病毒(Mesles virus,MV)属于副粘病毒科,是一种有包膜的病毒,具有不分节段的负链RNA,RNA基因组包含6个编码8种病毒蛋白的基因(图10)。该病毒在宿主细胞上有3个受体:信号淋巴细胞激活分子(SLAM/CD150)、CD46和nectin-4(脊髓灰质炎病毒受体样4,PVRL4)。由于CD46在许多癌细胞中的过度表达,MV具有感染肿瘤细胞的天然能力。除了自然感染机制外,一些人为修饰可以提高麻疹的溶瘤功效(图11),例如制造更强的癌症特异性嗜性、武装以提高癌细胞杀伤作用以及屏蔽以避免阻碍治疗功效的抗病毒免疫反应。由于麻疹疫苗的广泛使用,抗病毒免疫对于基于MV的溶瘤药物来说是一个特别困难的障碍。可用来避免脱靶副作用和将MV特异性地重新定向到癌细胞的策略包括插入肿瘤特异性配体、插入整合素结合肽、插入单链T细胞受体和修饰包膜融合特性。此外,1型干扰素应答的缺陷是MV肿瘤选择性的另一个原因。
MV-NIS是一种表达碘化钠同向转运体的MV突变体,受感染的肿瘤细胞对I131的摄取增加,从而在肿瘤细胞中产生放射毒性;其顺利性和病毒复制的合适性已在复发性多发性骨髓瘤患者的I期试验中得到证明。
在瑞士,使用麻疹病毒Edmonston-Zagreb(MV-EZ)株进行了一项I期临床试验,MV-EZ在5名皮肤T细胞淋巴瘤(CTCL)患者接受IFN-α预处理后进行瘤内给药。MV-EZ治疗引起IFNγ产生增加和CD4/CD8比率降低,带来了更好的预后。进一步的研究证明了不同溶瘤MV菌株的合适性和顺利性。
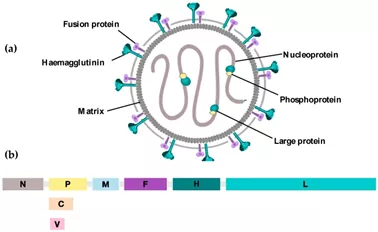
图10麻疹病毒及其基因组
(a)麻疹病毒的示意图。麻疹病毒是一种有包膜的负链RNA病毒。RNA基因组受核蛋白(N)保护,核蛋白(N)与RNA依赖性RNA聚合酶(RdRp)(称为大蛋白(L))及其辅因子磷蛋白(P)相关这些共同构成了被基质(M)包围的核糖核蛋白复合体(RNP)。两种病毒糖蛋白,血凝素(H)蛋白和融合(F)蛋白,从脂质双层突出并参与病毒进入宿主细胞。(b)麻疹病毒基因组。MV基因组由15,894个RNA核苷酸组成,包括六个转录单位,每个转录单位由三核基因间序列隔开。蛋白质V和C是由P基因的替代RNA转录物产生的非结构蛋白质。
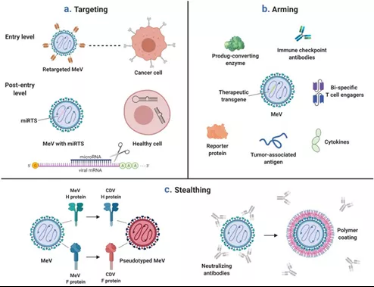
图11溶瘤MV的靶向、武装和隐身
(a)溶瘤MV的靶向性和肿瘤特异性可以在多个水平上进行设计。进入靶向:此处显示的是完全重新靶向的MV,它通过与MV H蛋白融合的scFv识别肿瘤抗原。进入后靶向:溶瘤MV携带微小RNA的靶位点,这些位点存在于健康细胞中,但在恶性细胞中丢失。这种microRNA控制的MV在表达同源microRNA的健康细胞中被强烈减弱,但对肿瘤细胞仍然完全合适。(b)溶瘤MV可以被设计编码治疗性转基因。(c)溶瘤MV的隐匿。左图:MV与密切相关的副粘病毒(犬瘟热病毒,CDV)的包膜糖蛋白的假型分析。右图:为了避免被预先存在的抗MV抗体中和,可以使用聚合物包膜结构屏蔽单个病毒粒子。
新城疫病毒
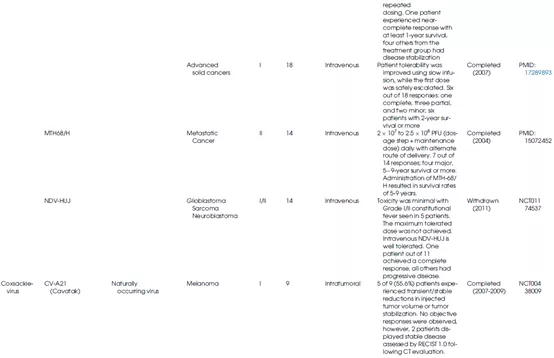
新城疫病毒(NDV)属于副粘病毒科,是一种具有非节段负链RNA的禽类包膜病毒,其血凝素-神经氨酸酶(HN)蛋白与宿主细胞上含唾液酸的受体结合会触发病毒的内吞作用。NDV的复制发生在细胞质中,且从未观察到与宿主基因组的任何重组。由于对IFN-α和IFN-β高度敏感,宿主防御机制会迅速阻止NDV复制,而癌细胞由于I型干扰素反应较弱,故而对NDV敏感(图7)。新城疫病毒的溶瘤活性与诱导癌细胞凋亡和通过增加细胞因子(IL-12、GM-CSF、RANTES和I型干扰素)激活固有免疫系统有关。研究表明,新城疫病毒HN蛋白是一种合适的抗原,能增强T淋巴细胞对肿瘤细胞的杀伤反应。此外,基因工程手段也可进一步改进NDV的溶瘤效果(图8)。
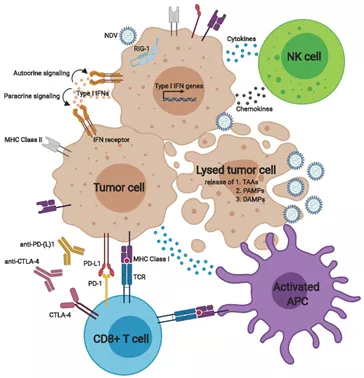
图7新城疫病毒(NDV)激活先天性和适应性抗肿瘤免疫反应
NDV选择性地感染具有缺陷的抗病毒防御的肿瘤细胞。由传感器(如RNA解旋酶RIG-1)介导的细胞外和细胞内信号传导导致I型IFN和相关基因的表达。自分泌和旁分泌IFN信号上调MHC I类和II类呈递、共刺激分子和细胞表面的免疫检查点。此外,细胞因子和趋化因子的释放募集了先天效应细胞,例如NK细胞和巨噬细胞以及抗原呈递细胞(APC)。病毒介导的直接溶瘤使得肿瘤抗原、PAMP和DAMP释放,激活APC。活化的APCs刺激T细胞,产生针对肿瘤和病毒抗原的细胞溶解性T细胞;然而,激活的T细胞的效应器功能可以通过上调肿瘤细胞和APCs上的PD-L1以及T细胞上的PD-1和CTLA-4而受到控制。这些负反馈机制的上调为新城疫病毒与免疫检查点控制剂的结合提供了理论基础。
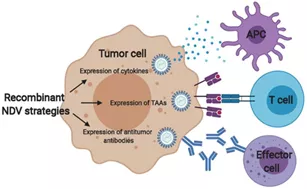
图8通过重组基因工程增强NDV抗肿瘤免疫反应
基因工程可用于产生具有更大潜力刺激抗肿瘤免疫反应的NDV毒株。首先,经过工程改造以表达细胞因子(例如GM-CSF或白细胞介素)的NDV可以增加先天效应细胞(例如抗原呈递细胞(APC))的募集。其次,NDV可用作靶向特定肿瘤抗原的治疗性疫苗,例如致癌病毒抗原、移码突变或突变的自身抗原。三,NDV可以被设计为表达单链可变片段或完整的抗肿瘤抗体,以通过效应细胞诱导抗体依赖性细胞毒性。
柯萨奇病毒
柯萨奇病毒属于小核糖核酸病毒科,是一种具有单链RNA基因组的无包膜病毒。与NDV一样,柯萨奇病毒在宿主细胞的细胞质中复制,降低了插入突变的可能性,并且不需要基因改造来实现溶瘤活性。
作为OV,研究较广泛的是柯萨奇病毒A21(CVA21),它是一种天然存在的肠道病毒。CVA21以细胞内粘附分子-1(ICAM-1)作为主要受体,利用衰变加速因子(DAF)作为辅助受体来感染宿主细胞。研究表明,一些实体瘤,包括黑色素瘤、结肠癌、乳腺癌、头颈癌、子宫内膜癌、肺癌和胰腺癌都高表达ICAM-1和DAF受体。因此,CVA21可能对癌细胞具有天然的趋向性。
柯萨奇病毒B3型(CVB3)是另一种重要的溶瘤柯萨奇病毒。研究表明,由于DAF和柯萨奇-腺病毒受体(CAR)的过度表达(图9),CVB3还具有对癌细胞的天然趋向性,是非小细胞肺癌细胞。CVB3的抗癌活性取决于其诱导细胞凋亡的能力。PI3K/AKT信号通路加速了CVB3的复制,由于该通路在大多数癌症中被激活,CVB3能够在这些癌细胞中选择性复制。
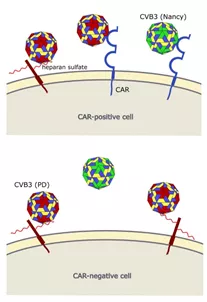
图9 CVB3与细胞受体的相互作用
原CVB3菌株Nancy使用CAR进入细胞,而CVB3变体PD可以通过N-和6-O-硫酸乙酰肝素和CAR感染细胞。上图:CAR阳性细胞可以感染PD和Nancy菌株。下图:当N-和6-O-硫酸乙酰肝素在细胞表面表达时,CAR阴性细胞不能被Nancy菌株感染,但会被PD感染。
其他病毒
除了上述病毒之外,还有许多其他病毒正在研究作为用于癌症治疗的溶瘤剂,包括脊髓灰质炎病毒、细小病毒、塞内卡谷病毒、水泡性口炎病毒、马拉巴病毒以及逆转录病毒。
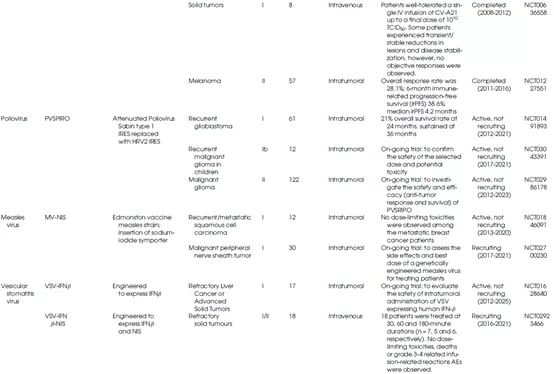
表1总结了不同种类的溶瘤病毒的生物学特性及其在临床上的应用情况。
表1 OV的生物学特性和临床应用
参考文献:
[1]Lan Q,Xia S,Wang Q,Xu W,Huang H,Jiang S,Lu L.Development of oncolytic virotherapy:from genetic modification to combination therapy.Front Med.2020Apr;14(2):160-184.doi:10.1007/s11684-020-0750-4.Epub 2020 Mar 7.PMID:32146606;PMCID:PMC7101593.
[1]Abd-Aziz N,Poh CL.Development of oncolytic viruses for cancer therapy.TranslRes.2021 Apr 24:S1931-5244(21)00104-3.doi:10.1016/j.trsl.2021.04.008.Epubahead of print.PMID:33905949.
[3]ChaurasiyaS,Fong Y,Warner SG.Oncolytic Virotherapy for Cancer:Clinical Experience.Biomedicines.2021 Apr 13;9(4):419.doi:10.3390/biomedicines9040419.PMID:33924556;PMCID:PMC8069290.
[5]Zhang Y,Li Y,Chen K,Qian L,Wang P.Oncolytic virotherapy reverses theimmunosuppressive tumor microenvironment and its potential in combination withimmunotherapy.Cancer Cell Int.2021 May 13;21(1):262.doi:10.1186/s12935-021-01972-2.PMID:33985527;PMCID:PMC8120729.
[6]Abudoureyimu M,Lai Y,Tian C,Wang T,Wang R,Chu X.Oncolytic Adenovirus-A Nova for Gene-Targeted Oncolytic Viral Therapy inHCC.Front Oncol.2019 Nov 8;9:1182.doi:10.3389/fonc.2019.01182.Erratum in:Front Oncol.2020 May 05;10:701.PMID:31781493;PMCID:PMC6857090.
[7]Menotti L,Avitabile E.Herpes Simplex VirusOncolytic Immunovirotherapy:The Blossoming Branch of Multimodal Therapy.Int JMol Sci.2020 Nov 5;21(21):8310.doi:10.3390/ijms21218310.PMID:33167582;PMCID:PMC7664223.
[9]Yang X,Huang B,Deng L,Hu Z.Progress in genetherapy using oncolytic vaccinia virus as vectors.J Cancer Res Clin Oncol.2018 Dec;144(12):2433-2440.doi:10.1007/s00432-018-2762-x.Epub 2018 Oct 6.PMID:30293118.
[10]Burman B,Pesci G,Zamarin D.NewcastleDisease Virus at the Forefront of Cancer Immunotherapy.Cancers(Basel).2020Nov 28;12(12):3552.doi:10.3390/cancers12123552.PMID:33260685;PMCID:PMC7761210.
[11]Geisler A,Hazini A,Heimann L,Kurreck J,Fechner H.Coxsackievirus B3-Its Potential as an Oncolytic Virus.Viruses.2021Apr 21;13(5):718.doi:10.3390/v13050718.PMID:33919076;PMCID:PMC8143167.
[12]Aref S,Bailey K,Fielding A.Measles to theRescue:A Review of Oncolytic Measles Virus.Viruses.2016 Oct 22;8(10):294.doi:10.3390/v8100294.PMID:27782084;PMCID:PMC5086626.
[13]Leber MF,Neault S,Jirovec E,Barkley R,SaidA,Bell JC,Ungerechts G.Engineering and combining oncolytic measles virus forcancer therapy.Cytokine Growth Factor Rev.2020 Dec;56:39-48.doi:10.1016/j.cytogfr.2020.07.005.Epub 2020 Jul 3.PMID:32718830;PMCID:PMC7333629.
[14]Müller L,Berkeley R,Barr T,Ilett E,Errington-Mais F.Past,Present and Future ofOncolytic Reovirus.Cancers(Basel).2020 Oct 31;12(11):3219.doi:10.3390/cancers12113219.PMID:33142841;PMCID:PMC7693452.

- 所属栏目:胶质瘤
- 如想转载“首款脑胶质瘤溶瘤病毒产品正式上市”请务必注明来源和链接。
- 网址:https://www.incsg.com/jiaozhiliu/1278.html
- 更新时间:2021-11-12 11:43:41




